Most amyloids1 are misfolded proteins, having enormous variety in native structures. Pathological amyloids are implicated in diseases including Alzheimer’s disease and many others. They are characterized by long, unbranched fibrillar structure, enhanced birefringence on binding Congo red dye, and cross-β structure – β-strands running approximately perpendicular to the fibril axis, forming long β-sheets running in the direction of the axis. Fiber diffraction patterns from amyloids are marked by strong intensity at about 4.8 Å in the meridional direction (parallel to the fibril axis), corresponding to the separation of strands in a β-sheet, and in many cases broader but distinct equatorial intensity at about 10 Å. The 10 Å intensity (whose position may vary considerably) comes from the distance between stacked β-sheets. This stacking is characteristic of the many amyloids formed by small peptides, including peptide fragments of larger amyloidogenic proteins. While some authors have required the 10 Å intensity to characterize an amyloid, it is not strictly necessary, since architecturally more complex examples have been found of Congo-red-staining fibrils with cross-β structure, but without the stacked-sheet structure, and consequently without the 10 Å intensity on the equator.
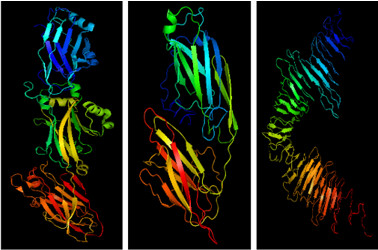
C123 (left), AgA (center), and Smu_63c (right).
Amyloids do not always stem from protein misfolding. Organisms across all kingdoms utilize functional amyloids in numerous biological processes. Bacteria are no exception. Bacterial amyloids contribute to biofilm formation and stability. Tooth decay is the most common infectious disease in the world. A major etiologic agent, Streptococcus mutans, is a quintessential biofilm dweller that produces at least three different amyloid-forming proteins, adhesins P1 and WapP, and the cell density and competence regulator Smu_63c2. The naturally occurring truncation derivatives of P1 and WapA, C123 and AgA, represent the amyloidogenic moieties, and a new paradigm of Gram-positive bacterial adhesins is emerging of adhesins having dual functions in monomeric and amyloid forms. While each S. mutans protein possesses considerable β-sheet structure, the tertiary structures of each protein are quite different (Fig. 1). This study further characterized S. mutans amyloids and addressed the ongoing debate regarding the underlying structure and assembly of bacterial amyloids including speculation that they are structurally dissimilar from better-characterized amyloids.

The highlighted work utilized classical x-ray fiber diffraction performed at the SSRL biological SAXS Beam Line 4-2 to characterize S. mutans amyloids that display an incompletely understood mat-like structure. During the course of sample preparation and protease digestion to remove residual monomers from amyloid preparations of purified proteins, it became apparent that typical amyloid fibers were visualized after proteolysis, and that mats were reconstituted by incubation of monomers with fibers. When microbiologists at the University of Florida were introduced by SSRL Beam Line 4-2 staff scientists to structural biologists from Vanderbilt University, the question of S. mutans basic amyloid structure was answered. In a series of synchrotron based fiber diffraction experiments at SSRL Beam Line 4-2, the research team established for the first time that the classical β-amyloid structure was indeed the core building block of both amyloid mats and fibers for all three bacterial amyloid-forming proteins. AgA is shown here as an example (Fig. 2).
The establishment of a conserved fiber diffraction pattern of S. mutans amyloids will enable future studies to evaluate biofilm matrix material of this and other microbes. This method and structural foundation can now be employed to assess the impact of varying environmental parameters on amyloid assembly within biofilms, and of potential treatment of biofilm-dependent diseases by amyloid inhibitors. Sometimes old-school is the most valuable tool!
- G. Stubbs and J. Stöhr, “Structural Biology of PrP Prions”, Cold Spring Harb. Perspect. Med. 7, 113 (2017)
- R. N. Besingi, I. B. Wenderska, D. B. Senadheera, D. G. Cvitkovitch, J. R. Long, Z. T. Wen, L. J. Brady, “Functional Amyloids in Streptococcus mutans, Their Use as Targets of Biofilm Inhibition and Initial Characterization of SMU_63c.”, Microbiology 163, 488 (2017)