
3-Hydroxyanthranilate-3,4-dioxygenase (HAO) is a critical, non-heme iron enzyme involved in tryptophan-kynurenine metabolism and NAD+ biosynthesis; it activates molecular oxygen to cleave an aromatic substrate. Its unstable, acyclic product, 2-amino-3-carboxymuconic semialdehyde (ACMS), is either metabolized to acetoacetate through a series of downstream enzymes, or auto-cyclizes to quinolinic acid (QUIN). All kingdoms of life use QUIN as a universal precursor for de novo biosynthesis of NAD+, and in mammals specifically, QUIN is formed by the non-enzymatic decay of the product of the HAO-catalyzed reaction. Overproduction of QUIN is associated with neurodegenerative and other undesired diseases. As a result, HAO has long been recognized as a potential drug target.1 Despite the early discovery and biological significance of HAO, its catalytic mechanism and how it balances downstream catalytic biomachinery versus non-enzymatic QUIN production has remained elusive.
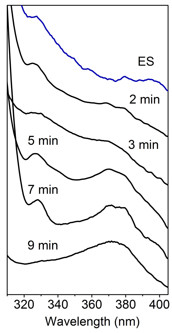
Major hindrances to further understand the catalytic and regulatory mechanism of HAO are its fast turnover (25 s-1), lack of intermediates, and undefined product conformation. The exact conformation of ACMS has not been characterized due to its short lifetime and a large number of possible conformers. To overcome these challenges, the highlighted work combined single-crystal spectroscopies and x-ray crystallography to probe changes in the active site and the overall conformation of HAO during in crystallo catalysis. The enzyme was first crystallized and then incubated with its aromatic substrate under anaerobic conditions. Subsequently, crystals of enzyme-substrate complex were exposed to oxygen for different times to interrogate any possible intermediates. The accumulation of catalytic intermediate in the crystalline state was identified using the integrated single-crystal UV-vis spectroscopy system at SSRL BL9-2. As shown in Fig. 1, substrate-bound HAO crystals exhibited a λmax at 327 nm. Upon exposure to oxygen for 2, 3, 5, 7, 9 min or longer exhibited features distinct from those of the substrate-enzyme complex, suggesting specific intermediates populate at these time points. Lastly, substrate-bound crystals mostly converted to product-bound form with a λmax at 374 nm. This result indicated that the crystals are catalytically active and the reaction rate in the crystalline state is more than 10,000-fold slower than in solution. More importantly, the considerably slower catalysis and evidence of accumulating intermediates provide a promising platform to further structurally characterize the reaction intermediates and enzyme-bound product. Furthermore, investigations of in crystallo HAO reaction using single-crystal electron paramagnetic resonance spectroscopy also support significantly slower turnover and detects the presence of a radical intermediate during catalysis.
Next, x-ray diffraction was employed to determine the atomic structure of each intermediate (confirmed by in-situ UV-vis spectroscopy) as well as associated active site loop movements.2 Besides substrate monodentate and bidentate bound structures, structures of intermediates after oxygen activation were also identified and characterized, including superoxo, alkylperoxo, seven-membered lactone, and two enol tautomers of the unstable dioxygenase product (Fig. 2). The seven-membered, mono-oxygenated lactone intermediate presumably corresponds to the radical species detected by EPR spectroscopy and supports a catalytic mechanism with stepwise incorporation of oxygen, providing a utility for future medicinal chemistry efforts. The finding of two enol tautomers unveils an unexpected isomerase activity upon product formation, which helps to understand how a metabolic enzyme competes with a non-enzymatic reaction to control the product outcome for distinct biological needs. A total of 7 high-resolution intermediate structures allows for a step-by-step visualization of the catalytic cycle and protein dynamics during catalysis (see Enzyme in Action Movie in PNAS Figures & SI). Overall, these results reveal a detailed dioxygenase mechanism along with potential isomerization activity that fine-tunes product profiling and affects the production of QUIN at a junction of the metabolic pathway.
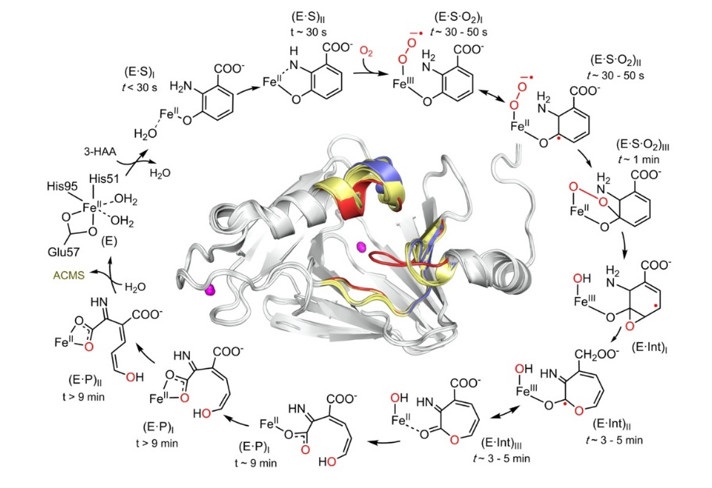
Figure 2. HAO catalytic cycle defined by the characterized intermediates.
- T. W. Stone and L. G. Darlington, “Endogenous Kynurenines as Targets for Drug Discovery and Development”, Nat. Rev. Drug Discov. 1, 609 (2002).
- Y. Yang, F. Liu and A. Liu. “Adapting to Oxygen: 3-Hydroxyanthranilate 3,4-dioxygenase Employs Loop Dynamics to Accommodate Two Substrates with Disparate Polarities”, J. Biol. Chem., 293, 10415 (2018)
Y. Wang, K. F. Liu, Y. Yang, I. Davis and A. Liu, “Observing 3-hydroxyanthranilate-3,4-dioxygenase in Action through a Crystalline Lens”, Proc. Natl. Acad. Sci. U. S. A. 117, 19720 (2020) doi: 10.1073/pnas.2005327117
