
T cells are tasked with detecting and counteracting infection and cellular dysfunction within an organism. Each T cell expresses a unique T-cell receptor (TCR), which is selected during T-cell development to bind to major histocompatibility complexes (MHCs) that display short (~8-15 amino acid) linear peptide antigens. The expanding compendium of TCR-peptide-MHC (TCR-pMHC) complex structures has revealed that essentially all TCR-pMHC interactions follow general rules, where the germline-encoded TCR CDR1 and CDR2 loops primarily contact the MHC helices while the diverse TCR CDR3 loops contact the peptide (1-2).
Similar to antibodies, the TCR repertoire contains a huge degree of combinatorial diversity to ensure that any potential antigen can be detected. Despite this diversity, the TCR repertoire faces a numbers problem – the number of possible peptide antigens displayed by MHC dwarfs the total number of T cells in a given organism. Since there are no systematic ‘holes’ in the immune repertoire, each TCR must be able to recognize a large number of peptides. While a need for TCR cross-reactivity has previously been articulated (3), the tools have not existed to experimentally quantitate the degree and structural mechanism of cross-reactivity.
To address this question, the researchers combined a bevy of combinatorial, structural, and computational approaches that allowed to comprehensively capture the peptide repertoire for TCRs of interest. They selected highly diverse peptide-MHC libraries displayed on yeast cells with five multimerized TCRs and then sequenced all the clones that enriched from the libraries. Surprisingly, it was found that in all cases, the recovered peptide sequences had readily apparent homology with the previously known ‘wild-type’ peptide ligands.
In addition to wild-type-like peptides, several of the TCRs also selected peptides with seemingly little homology to the dominant TCR contact motif, sharing as few as one out of five TCR contacts. To determine how the TCRs were able to recognize such different peptides, three TCR-pMHC complexes containing these alternate peptides were crystallized and x-ray diffraction data were measured using Beam Lines 11-1 and 12-2 at the Stanford Synchrotron Radiation Lightsource (SSRL). Use of the Stanford Automated Mounting system and a Pilatus detector were crucial to obtain good quality data to 2.6-3.3 Å resolution.
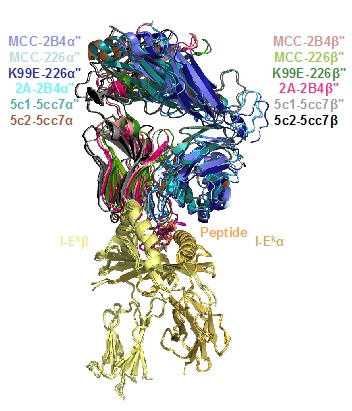
When compared with the previously solved ‘wild-type’ TCR-pMHC complexes (4), the solved structures retain the previously observed TCR-pMHC ‘docking’ footprint and the same TCR CDR1 and CDR2 contacts (Figure 1). The TCR CDR3 loops reorient to accommodate the alternate peptide residues. These structural relationships could not have been easily predicted through examining the peptide sequences alone. When the structural information was combined with sequence clustering of all selected peptides for each given TCR, it was found that essentially all peptides selected by each TCR comprise a single sequence family, showing linkages through both sequence and structure.
These results show that TCR cross-reactivity can be defined in two complementary ways. Each TCR is likely able to recognize thousands of peptides, satisfying the requirement to be able to recognize peptide antigens from a wide array of pathogens. Structurally, TCR cross-reactivity appears to manifest itself though relatively conservative substitutions to the TCR recognition epitope of the peptide with a greater degree of substitution allowed away from the central TCR-peptide interaction site. While there are likely to be exceptions to these rules, this mechanism allows the TCR to recognize a limited number of unique epitopes spanning a large number of unique peptides, satisfying the need for cross-reactivity while avoiding degenerate recognition.
With this knowledge in hand, the researchers have been able to leverage the limited cross-reactivity of TCRs to search for peptide ligands for TCRs of interest that do not exist in our libraries. Since the number of possible peptide ligands greatly surpasses what can be assayed in a yeast display library, the chance that any given peptide of interest will be found is low. However, since the peptides that are found in the library are likely to be related to a biological peptide of interest, it has now been shown that it is possible to find TCR agonist peptides present in pathogens, the environment, and even the host organism itself with a high success rate. Together, these findings suggest a way forward to find disease-relevant T-cell ligands in contexts where few if any are known, such as in many types of cancer and autoimmunity.
- K. C. Garcia and E. J. Adams, "How the T Cell Receptor Sees Antigen--a Structural View", Cell 122, 333 (2005)
- M. G. Rudolph, R. L. Stanfield, and I. A. Wilson, "How TCRs Bind MHCs, Peptides, and Coreceptors", Ann. Rev. Immunol. 24, 419 (2006)
- D. Mason, "A Very High Level of Crossreactivity is an Essential Feature of the T-cell Receptor", Immunol. Today 19, 395 (1998)
- E. W. Newell, L. K. Ely, A. C. Kruse, P. A. Reay, S. N. Rodriguez, A. E. Lin, M. S. Kuhns, K. C. Garcia, and M. M. Davis, "Structural Basis of Specificity and Cross- Reactivity in T Cell Receptors Specific for Cytochrome c-I-Ek", J. Immunol. 186, 5823 (2011)
M. E. Birnbaum, J. L. Mendoza, D. K. Sethi, S. Dong, J. Glanville, J. Dobbins, E. Özkan, M. M. Davis, K. W. Wucherpfennig and K. C. Garcia, "Deconstructing the Peptide-MHC Specificity of T Cell Recognition", Cell 157, 1073 (2014), DOI: 10.1016/j.cell.2014.03.047
