| SSRL Science Highlight - January 2008 | |

| ||||||||||||||
|
Proteins are delivered to specific sites within cells in small
membrane-enclosed carriers called transport vesicles. Transport vesicles are
targeted to the correct acceptor membrane by a number of sequential steps that
are regulated by small GTPases of the Rab and Arf families. Small GTP-binding
proteins (GTPases) are a large group of proteins involved in the regulation of
quite different cellular processes like cell proliferation, differentiation
(Ras-, Rap- and Ral-family), nuclear transport (Ran), vesicular transport
(Rab-family) and cytoskeleton organization (Rho- and Arf-family). Vesicles are
transported along microtubule or actin tracks; target recognition is thought to
involve a molecular "tethering" event at the target membrane that is mediated
by coiled-coil and multi-subunit tethers, prior to membrane fusion.
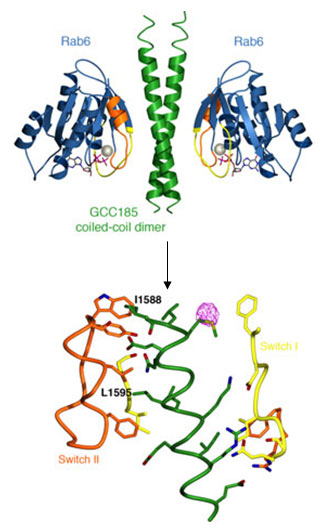
Figure 1. (Top) Ribbon representation of the Rab6-GCC185 complex. Switch I (yellow)
and II (orange) regions in Rab6 (blue) bind the GCC185 Rab binding domain
(green). Rab6-bound GTP (stick model) and magnesium (sphere) are shown.
(Bottom) View of the Rab6-GCC185 binding interface. Residues in GCC185 that are critical
for Rab6 interaction and Golgi localization are labeled. An anomalous
difference Fourier density map (pink) indicates the position of a
selenomethionine residue used to build the GCC185 model.
Given that GCC185 plays vital roles both in organizing the cell cytoskeleton
and in vesicle traffic, Schweizer Burguete et al. investigated how the putative
tether itself is localized to the Golgi membrane. We have shown that
Golgi-recruitment of GCC185 is mediated by the cooperation of two
Golgi-localized small GTPases belonging to the Rab and Arf families. Rab6
binding to GCC185 promotes the subsequent binding of Arl1 to an immediately
adjacent domain. Biochemical analysis of these interactions revealed a
helical, dimeric, Rab6 binding domain in GCC185. The crystal structure of a
complex between the Rab binding domain (RBD) of GCC185 and Rab6 was determined using
diffraction data obtained at SSRL (Beamlines 11-1 and 7-1) and provided the
stoichiometry and the molecular details of this interaction. Rab6 switch I and
II regions, which adopt a specific conformation when the protein is GTP-bound,
contact a dimeric coiled-coil in GCC185 with two-fold symmetry, and residues
critical for Rab binding and Golgi-localization of GCC185 lie in the binding
interface (Figure 1).
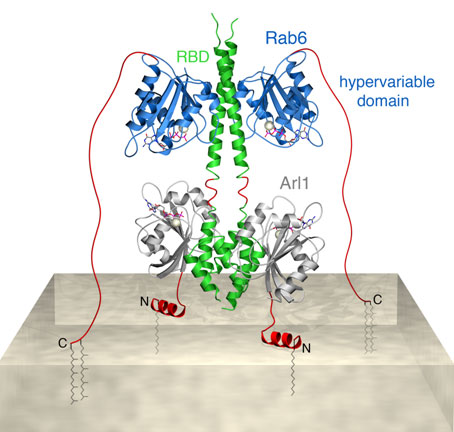
Based on our observations we have created a structure-derived model for
simultaneous GTPase binding to the carboxy-terminal region of GCC185. In this
model the three proteins form a hetero-hexameric complex that attach the 185
kDa tether to the surface of Golgi membranes (Figure 2). This model highlights
how Arf and Rab-family members may interact with the same binding partner at
different distances from the membrane. The Rab GTPases are expected to reach
binding sites as far as 10 nm away from the membrane via their unstructured and
membrane-anchored, C-terminal tails. Arf GTPases on the other hand, will bind
to membrane-proximal domains, enabling cooperation with Rab proteins in
determining the fate of a common binding partner.
Figure 2. Model for dual GTPase binding to adjacent RBD and GRIP domains
in GCC185. Rab6 (blue) and Arl1 (gray) anchor GCC185 (green) to Golgi membranes
by inserting prenyl and myristoyl groups (gray stick models) respectively into
the cytosolic leaflet of the lipid bilayer (beige). This model was generated by
combining the Rab6-GCC185 crystal structure with that of a modeled Arl1-GCC185
GRIP domain complex. Regions absent from the two crystal structure models are
shown in red.
Primary Citation
References
Efimov, A., Kharitonov, A., Efimova, N., Loncarek, J., Miller, P. M.,
Andreyeva, N., Gleeson, P., Galjart, N., Maia, A. R., McLeod, I. X., Yates, J.
R., Maiato, H., Khodjakov, A., Akhmanova, A. and Kaverina, I. (2007).
Asymmetric CLASP-dependent nucleation of noncentrosomal microtubules at the
trans-Golgi network. Dev Cell. 12(6):917-30.
Panic, B., Perisic, O., Veprintsev, D. B., Williams, R. L. and Munro, S. (2003)
Structural basis for Arl1-dependent targeting of homodimeric GRIP domains to
the Golgi apparatus. Mol Cell. 12(4):863-74.
|
| Last Updated: | 23 January 2008 |
| Content Owner: | A. Schweizer Burguete, T.D. Fenn, A.T. Brunger and S.R. Pfeffer |
| Page Editor: | L. Dunn |