 Sompop Bencharit1,2 and Matthew R. Redinbo1,3
Sompop Bencharit1,2 and Matthew R. Redinbo1,3
1Department of Chemistry, University of North Carolina at Chapel
Hill, Chapel Hill, North Carolina
2School of Dentistry, University of North Carolina at Chapel Hill,
Chapel Hill, North Carolina
3Department of Biochemistry & Biophysics and the Lineberger
Comprehensive Cancer Center, University of North Carolina at Chapel Hill,
Chapel Hill, North Carolina
 | |
| Figure 1
A proposed mechanism for the activation of irinotecan (CPT-11) by rCE. CPT-11 (orange) enters from the top of the catalytic gorge and fits well into the active site (catalytic Ser-221 and Glu-353 in green). After cleavage, the alcohol product (SN-38; magenta) leaves via the catalytic gorge, while the acyl product (4PP; purple) moves past the gate residues (cyan) and docks adjacent to the regulatory domain (red) on the surface of the molecule. |
|
First, the structure of a mammalian carboxylesterase, that of rabbit liver carboxylesterase (rCE), reveals the activation mechanism of the camptothecin anticancer drug, irinotecan (CPT-11) (Bencharit et al., 2002). Using the position of a bound product on the surface of the enzyme, we have proposed that rCE utilizes a "side door" secondary product exit pore to facilitate the efficient processing of irinotecan (Fig 1).
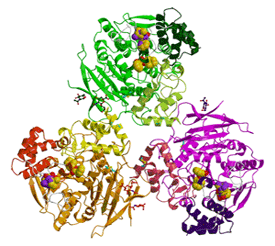
Second, our work on human carboxylesterase 1(hCE1), the human homolog of rCE, are the first structures of a human enzyme bound to cocaine and heroin analogues (Fig 2a-b) (Bencharit et al., 2003a). The hCE1 active site contains both specific and promiscuous compartments, which enable the enzyme to act on structurally distinct chemicals. The position of bound homatropine, a cocaine analogue, and bound naloxone, a heroin analogue, reveal how hCE1 utilizes its large flexible pocket promiscuously and its small rigid pocket specifically. These crystal structures together with the results from atomic force microscopy provide the first evidence of hCE1's trimer-hexamer equilibrium.
 | 
| |||
| Figure 2
a. The hCE1 trimer as observed in the homatropine complex; homatropine molecules bound at the active sites are gold. Homatropines bound at the Z site, between the Ω loops, are gold and purple. b. Two orientations of naloxone methiodide (green and orange) bound within the hCE1 catalytic pocket. The two binding modes of this heroin analog are related by an opposite two-fold rotation about the vertical axis. The molecular surfaces of the catalytic binding pocket corresponding to the two orientations of the naloxone ligand are rendered in transparent green and orange, and demonstrate the induced fit property of the substrate-binding pocket. | ||||
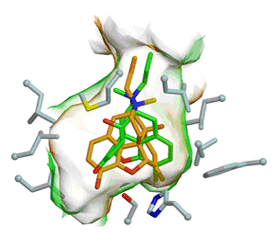
 | |
| Figure 3 2.4 Ĺ resolution simulated annealing omit maps (contoured at 3.0s) showing the electron density for tacrine bound at the active site of hCE1. Multiple conformations of the ligand fitted into the density are shown in different colors. |
Third, we have presented the structure of hCE1 in complexes with an anti-Alzheimer drug tacrine (Bencharit et al., 2003b). The manner in which tacrine binds, and the shape and size of hCE1 substrate binding gorge, are distinct from the tacrine bound acetylcholinesterase structure determined previously. Furthermore, tacrine appears in multiple conformations within hCE1's substrate binding gorge. We conclude that hCE1's ability to bind the same ligand in up to five conformations may contribute to its promiscuity and facilitate its ability to process variety of endogenous and exogenous compounds (Fig 3).
Diffraction data for the rCE, hCE1-homatropine, and hCE1-tacrine structures were collected at SSRL Beam Lines 7-1, 9-1, and 9-2. Diffraction data for the hCE1-naloxone structure was collected at the Advanced Photon Source (APS); SER-CAT Beam Line 22-ID.
References
- Redinbo MR, Bencharit S, Potter PM. Human carboxylesterase 1: from drug metabolism to drug discovery. Biochem. Soc. Trans. 2003;31:620-4.
- Bencharit S, Morton CL, Howard-Williams EL, Danks MK, Potter PM, Redinbo MR. Structural insights into CPT-11 activation by mammalian carboxylesterases. Nat. Struct. Biol. 2002;9:337-42.
- Bencharit S, Morton CL, Xue Y, Potter PM, Redinbo MR. Structural basis of heroin and cocaine metabolism by a promiscuous human drug-processing enzyme. Nat. Struct. Biol. 2003a;10:349-56.
- Bencharit S, Morton CL, Hyatt JL, Kuhn P, Danks MK, Potter PM, Redinbo MR. Crystal structure of human carboxylesterase 1 complexed with the Alzheimer's drug tacrine. From binding promiscuity to selective inhibition. Chem. Biol. 2003b;10:341-9.
SSRL Highlights Archive