
In the present study, X-ray absorption spectroscopy and Mössbauer spectroscopy
were used to identify the structural features consistent with the mechanistic
model, i.e. the presence of a short Fe(IV)-oxo interaction and a metal bound
halide. The Mössbauer spectra collected prior to XAS data collection
demonstrated that the quenched samples contained ~80% of the Fe(IV)
intermediate. Our XAS spectrum of the intermediate showed a large enhancement
in the 1s->3d transition pre-edge intensity relative to the anaerobic, reduced
control, which is consistent with the presence of an asymmetrical Fe(IV)=
O2- unit. The XANES edge energy is consistent with the 80% Fe(IV):
20% Fe(II) composition determined using Mössbauer spectroscopy. The most
compelling evidence for the presence of a formal Fe(IV)=O2- species
comes from the fitting analysis of the EXAFS oscillations. Fits to the
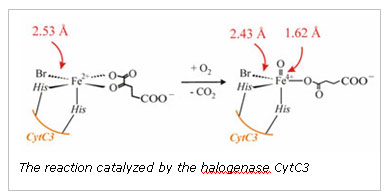
Fourier-filtered data require a short, 1.62 (± 0.02) Ĺ Fe-O interaction to best
model the data. Furthermore, a large scatterer is apparent in the Fourier
transformation of the data. This peak can be modeled with an Fe-Br interaction
at 2.43 Ĺ. If the coordination number of the short Fe-O interaction and the
Fe-Br interactions are systematically varied, the optimal coordination number
is 0.7-0.8 for both features, matching the sample composition determined by
Mössbauer spectroscopy. In contrast, fits to the reduced control sample were
not improved by adding a short Fe-O interaction. The Fe-Br interaction in the
control is 2.53 Ĺ, consistent with the distance found in the crystal structure
of SyrB2. The structural features we identified using XAS are only consistent
with a Br-Fe(IV)=O2- unit and confirms a key component of the proposed
mechanism.
Primary Citation
References
There are over 4,500 known halogenated natural products. The presence of a
halogen in the molecular framework tunes a compound's chemical reactivity or
biological activity in these natural fungicides and antibiotics. Four classes
of enzymes are now known to catalyze halogenation reactions: 1) vanadium
haloperoxidases, 2) heme haloperoxidases, 3) flavin-dependent halogenases, and
4) non-heme iron, alpha-ketoglutarate (aKG) dependent
halogenases.1,2 Walsh and
coworkers first identified the
aKG dependent
oxygenases in 20053 and noted that they catalyze the insertion of
halides into unreactive substrates (for example, the chlorination of a terminal
methyl group in barbamide4). The chemical logic of the non-heme
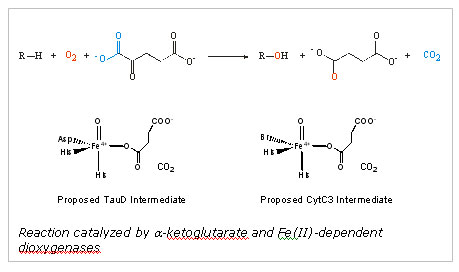
iron halogenases follows that of the aKG dependent,
non-heme iron dioxygenases such as TauD, the E. coli enzyme catalyzing
the hydroxylation of taurine (2-amino-1-ethanesulfonic acid).
5,6
For TauD, a high valent Fe(IV)-oxo species is generated that abstracts a
hydrogen atom from the substrate and a subsequent radical rebound results in
the hydroxylation of the substrate.7-9 A conserved His-His-Asp/Glu
"facial triad" provides the iron ligands.
Key insight into the mechanistic differences and similarities of the
aKG-dependent halogenases and oxygenases came from
the crystal structure of the halogenase SyrB2, responsible for the
chlorination reaction in the synthesis of syringomycin, a compound secreted by
Pseudomonas syringae.10 The structure indicated that a
halogen (Br or Cl) directly binds to the iron in place of the conserved
Asp/Glu. Thus, when a ferryl-oxo intermediate is generated upon
decarboxylation of the bound aKG, the intermediate
abstracts a hydrogen atom from the substrate and a halogen radical is
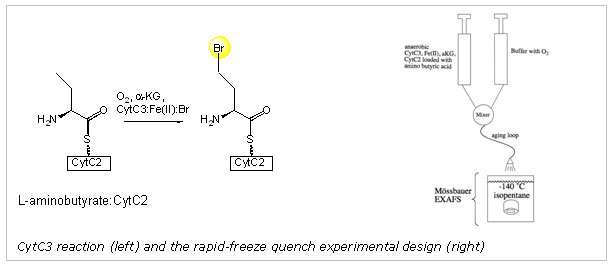
transferred to the substrate radical. Using the halogenase CytC3, an enzyme
capable of halogenating L-aminobutyrate, the Bollinger, Krebs, and Walsh groups
verified a key component of these proposed mechanistic steps. They
demonstrated that a high valent iron intermediate accumulates when the
halogenase CytC3 is exposed to oxygen in the presence of Cl-, aKG, and the
scaffold enzyme CytC2-substrate complex.11 They furthermore demonstrated that
this Fe(IV) species was indeed responsible for hydrogen atom abstraction from
the substrate.
Galonic Fujimori, D., Barr, E. W., Matthews, M.L., Koch, G. L., Yonce, J. R.*,
Walsh, C. T., Bollinger, J. M., Jr., Krebs, C., Riggs-Gelasco, P. J.
"Spectroscopic Evidence for a High-Spin Br-Fe(IV)-Oxo Intermediate in the
a-Ketoglutarate-Dependent Halogenase CytC3 from
Streptomyces", 2007, J. Am.
Chem. Soc., 129, 13408-13409.
SSRL is supported by the Department of Energy, Office of Basic Energy Sciences. The SSRL Structural Molecular Biology Program is supported by the Department of Energy, Office of Biological and Environmental Research, and by the National Institutes of Health, National Center for Research Resources, Biomedical Technology Program, and the National Institute of General Medical Sciences.