 Though life on earth is composed of a diverse range of organisms, some with
many different types of tissues and cells, all these are encoded by a molecule
we call DNA. The information required to build a protein is stored in DNA
within the cells. Not all the message in the DNA is used in each cell and not
all the message is used all the time. During cell differentiation, the cells
become dedicated for their specific function which involves selectively
activating some genes and repressing others. Gene regulation is an important
event in the developmental biology and the biology of various diseases, but a
more complex process.
Though life on earth is composed of a diverse range of organisms, some with
many different types of tissues and cells, all these are encoded by a molecule
we call DNA. The information required to build a protein is stored in DNA
within the cells. Not all the message in the DNA is used in each cell and not
all the message is used all the time. During cell differentiation, the cells
become dedicated for their specific function which involves selectively
activating some genes and repressing others. Gene regulation is an important
event in the developmental biology and the biology of various diseases, but a
more complex process.
In the bacteria there are distinct enzymes while one is capable of cleaving
DNA, the other protects DNA by modification. The complementary function
provided by the set of enzymes offers a defense mechanism against the phage
infection and DNA invasion. The incoming DNA is cleaved sequence specifically
by the class of enzymes called restriction endonuclease (REase). The host DNA
is protected by the sequence specific action of matching set of enzymes called
the DNA methyltransferase (MTase). The control of the relative activities of
the REase and MTase is critical because a reduced ratio of MTase/REase activity
would lead to cell death via autorestriction. However too high a ratio would
fail to provide protection against invading viral DNA. In addition a separate
group of proteins capable of controlling R-M proteins have been identified in
various restriction-modification (R-M) systems which are called C proteins
(Roberts et al., 2003).
A homolog of R-M C protein, named as C.BclI regulates the expression level of
M.BclI (methyltransferase) in E. coli differently. In the absence of
C.BclI, M.BclI was over-expressed and displayed relaxed specificity for
methylation of non-cognate (non-specific or less specific) sites. However, in
the presence of C.BclI and C box (the C.BclI binding sequence), the expression
level of M.BclI was down regulated and M.BclI did not show relaxed specificity.
| |
 |
|
Figure 1
Dimer of C.BclI structure. Two molecules that form the dimer are related by a
2-fold rotational symmetry. There are five alpha helices (A, B, C, D, E) in
each monomer of C.BclI and of them HTH motif is formed by the helices B and C.
|
Balendiran's group at City of Hope collected the x-ray diffraction data on
BL1-5 at SSRL. The crystal structure of C.Bcll was determined to 1.8 Å
resolution by anomalous dispersion methods, using mercury derivatives. The
high-resolution crystal structure of C.BclI uncovers the presence of a
helix-turn-helix (HTH) motif (Figure 1) and the potential to bend B-DNA around
the C.BclI dimer (Figure 2). Furthermore interactions with DNA are likely
mediated by residues on the surface of the HTH motif helices between 34 and 48
in the structure. Moreover by analogy to known proteins with HTH motif
(Hochschild et al., 1983; Bushman
and Ptashne, 1988; Bushman et al., 1989) highly conserved residue Glu27
in the C family of proteins and partly conserved Asp31 residue in C.BclI may
play a crucial role in the interaction with RNA polymerase and be important for
transcriptional control.
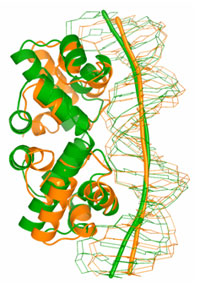 |
|
Figure 2
Comparison of C.BclI structure with the known structures containing HTH motif.
Induced DNA bending.
|
Primary Citation
Sawaya, M. R. Zhu, Z., Mersha, F., Chan, S-h., Dabur, R., Xu, S-y., Balendiran,
G. K., Crystal structure of the restriction-modification system control element
C.BclI and mapping of its binding site. Structure 13:1837-1847,
(2005).
Acknowledgement
The study was conducted in collaboration with Shuang-yong Xu, Ph.D., senior
scientist, New England Biolabs, with assistance from Professor David Eisenberg,
Professor Richard Dickerson, Duilio Cascio Ph.D., Michael R. Sawaya, Ph.D.,
University of California at Los Angeles; Stanford Synchrotron Radiation
Laboratory and SSRL staff; Jim D'Aoust, Project Manager, Cyberinfrastructure
Partnership, San Diego Supercomputer Center; Richard Roberts, Ph.D., CSO, Nobel
laureate, and Elisabeth Raleigh Ph.D., research director, New England Biolabs
Inc.
References
-
Bushman, F. D., and Ptashne, M. (1988). Turning lambda Cro into a
transcriptional activator. Cell 54, 191-197.
-
Bushman, F. D., Shang, C., and Ptashne, M. (1989). A single glutamic acid
residue plays a key role in the transcriptional activation function of lambda
repressor. Cell 58, 1163-1171.
-
Hochschild, A., Irwin, N., and Ptashne, M. (1983). Repressor structure and the
mechanism of positive control. Cell 32, 319-325.
-
Roberts, R. J., Belfort, M., Bestor, T., Bhagwat, A. S., Bickle, T. A.,
Bitinaite, J., Blumenthal, R. M., Degtyarev, S., Dryden, D. T., Dybvig, K.,
et al. (2003). A nomenclature for restriction enzymes, DNA methyltransferases,
homing endonucleases and their genes. Nucleic Acids Res 31,
1805-1812.
|
| PDF Version | | Lay Summary
|
| Highlights
Archive |
|
SSRL is supported by the Department of Energy, Office of Basic Energy Sciences.
The SSRL Structural Molecular Biology Program is supported by the Department of
Energy, Office of Biological and Environmental Research, and by the National
Institutes of Health, National Center for Research Resources, Biomedical
Technology Program, and the National Institute of General Medical Sciences.
|
|
|
